La traducción es el paso de la información transportada por el ARN-m a proteína.
La especificidad funcional de los polipéptidos reside en su secuencia lineal de aminoácidos que determina su estructura primaria, secundaria y terciaria. De manera, que los aminoácidos libres que hay en el citoplasma tienen que unirse para formar los polipéptidos y la secuencia lineal de aminoácidos de un polipéptido depende de la secuencia lineal de ribonucleótidos en el ARN que a su vez está determinada por la secuencia lineal de bases nitrogenadas en el ADN.
Los elementos que intervienen en el proceso de traducción son fundamentalmente: los aminoácidos, los ARN-t (ARN transferentes), los ribosomas, ARN-r (ARN ribosómico y proteínas ribosomales), el ARN-m (ARN mensajero), enzimas, factores proteicos y nucleótidos trifosfato (ATP, GTP).
El primer paso que tiene que producirse es la activación de los aminoácidos y formación de los complejos de transferencia. Los aminoácidos por sí solos no son capaces de reconocer los tripletes del ARN-m de manera que necesitan unirse a un ARN de pequeño tamaño (constante de sedimentación 4S) llamado ARN adaptador, ARN soluble o ARN transferente. Crick (1958) postuló la necesidad de la existencia de un adaptador que acoplará cada aminoácido a su correspondiente codón.
ESTRUCTURA DE LOS ARN TRANSFERENTES (ARN-t)
Los primeros estudios sobre la estructura de los ARN-t se realizaron por R. W. Holley y col. (1965) trabajando con el ARN-t de alanina de levaduras. A partir de sus 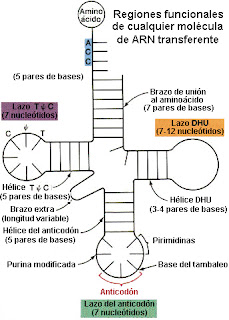 trabajos se estableció el modelo general de estructura de los ARN-t y por estas investigaciones recibió el Premio Nobel en (1968). Las moléculas encargadas de transportar los aminoácidos hasta el ribosoma y de reconocer los codones del ARN mensajero durante el proceso de traducción son los ARN transferentes (ARN-t). Los ARN-t tienen una estructura en forma de hoja de trébol con varios sitios funcionales:
trabajos se estableció el modelo general de estructura de los ARN-t y por estas investigaciones recibió el Premio Nobel en (1968). Las moléculas encargadas de transportar los aminoácidos hasta el ribosoma y de reconocer los codones del ARN mensajero durante el proceso de traducción son los ARN transferentes (ARN-t). Los ARN-t tienen una estructura en forma de hoja de trébol con varios sitios funcionales:
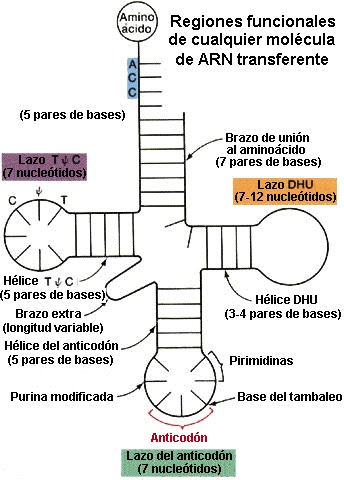 trabajos se estableció el modelo general de estructura de los ARN-t y por estas investigaciones recibió el Premio Nobel en (1968). Las moléculas encargadas de transportar los aminoácidos hasta el ribosoma y de reconocer los codones del ARN mensajero durante el proceso de traducción son los ARN transferentes (ARN-t). Los ARN-t tienen una estructura en forma de hoja de trébol con varios sitios funcionales:
trabajos se estableció el modelo general de estructura de los ARN-t y por estas investigaciones recibió el Premio Nobel en (1968). Las moléculas encargadas de transportar los aminoácidos hasta el ribosoma y de reconocer los codones del ARN mensajero durante el proceso de traducción son los ARN transferentes (ARN-t). Los ARN-t tienen una estructura en forma de hoja de trébol con varios sitios funcionales:- Extremo 3': lugar de unión al aminoácido (contiene siempre la secuencia ACC).
- Lazo dihidrouracilo (DHU): lugar de unión a la aminoacil ARN-t sintetasa o enzimas encargadas de unir un aminoácido a su correspondiente ARN-t.
- Lazo de T ψ C: lugar de enlace al ribosoma.
- Lazo del anticodón: lugar de reconocimiento de los codones del mensajero.
Normalmente el ARN-t adopta una estructura de hoja de trébol plegada en forma de L o forma de boomerang.
El que realiza el reconocimiento del codón correspondiente del ARN-m es el anticodón del ARN-t y no el aminoácido.
LOS RIBOSOMAS (ARN RIBOSÓMICO Y PROTEÍNAS RIBOSOMALES)
El reconocimiento entre los tripletes del mensajero y los anticodones de los ARN-t cargados con su  correspondiente aminoácido, así como el establecimiento de los enlaces peptídicos entre dos aminoácidos sucesivos tiene lugar en los ribosomas.
correspondiente aminoácido, así como el establecimiento de los enlaces peptídicos entre dos aminoácidos sucesivos tiene lugar en los ribosomas.
 correspondiente aminoácido, así como el establecimiento de los enlaces peptídicos entre dos aminoácidos sucesivos tiene lugar en los ribosomas.
correspondiente aminoácido, así como el establecimiento de los enlaces peptídicos entre dos aminoácidos sucesivos tiene lugar en los ribosomas.Los ribosomas son unas estructuras o partículas citoplásmicas formadas por ribonucleoproteínas (unión de ARN ribosómicos con proteínas ribosomales). Los ribosomas en las células eucarióticas se encuentran en la membrana del retículo endoplasmático. La estructura general de los ribosomas procarióticos y eucarióticos consta de una subunidad pequeña, una subunidad grande y dos sedes, la sede aminoacídica (Sede A) lugar de entrada de los ARN-t cargados con un aminoácido (aminoacil-ARN-t) y la sede peptídica (Sede P) lugar en el que se encuentran los ARN-t cargados con un péptido (peptidil-ARN-t).
ACTIVACIÓN DE LOS AMINOÁCIDOS Y FORMACIÓN DE LOS COMPLEJOS DE TRANSFERENCIA
La activación de los aminoácidos para formar los complejos de transferencia es el paso previo necesario para que pueda comenzar la traducción, y consiste en la unión de cada aminoácido a su ARN-t específico mediante la intervención de un enzima, la aminoacil-ARN-t sintetasa y el aporte de energía del ATP.aa1 + ARN-t1 + ATP → ARN-t1-aa1 + AMP + PPi
La unión del aminoácido al ARN-t tiene lugar por el extremo 3' del ARN-t. Todos los ARN-t en su extremo 3' contienen la secuencia 3' ACC 5'. Las aminoacil-ARN-t-sintetasas tienen tres sedes distintas, una para el reconocimiento del aminoácido, otra para el ARN-t y otra para el ATP. Debe existir al menos una aminoacil-ARN-t-sintetasa diferente por cada ARN-t distinto. El ARN-t se une a la aminoacil-ARN-t-sintetasa a través del lazo dihirouracilo (DHU).Por último, la especificidad de reconocimiento de las aminoacil-ARN-t-sintetasas y el correspondiente aminoácido no reside en el anticodón del ARN-t. Esta especificidad es lo que se ha llamado el Segundo Código Genético. Esta especificidad reside en el par de bases G y U que ocupan las posiciones 3 y 70, respectivamente del ARN-t. La ausencia de este par impide que se una la alanina a su ARN-t y la introducción de dicho par en la misma posición en los ARN-t-cys y ARN-t-phe les confiere la capacidad de unirse al aminoácido alanina.
INCORPORACIÓN DE LOS AMINOÁCIDOS A LA CADENA POLIPEPTÍDICA
Una vez activados los aminoácidos y formados los complejos de transferencia (ARN-t cargados con el aminoácido correspondiente) ya puede comenzar la síntesis de la cadena polipeptídica y la incorporación de los aminoácidos. En este proceso se pueden distinguir tres fases diferentes:

Elongación de la cadena polipeptídica.
Terminación de la cadena polipeptídica.
Iniciación

El proceso de iniciación de la traducción en las procariotas.La iniciación de la traducción en las procariotas supone ensamblar los componentes del sistema de traducción, que son: las dos subunidades ribosomales, el ARNm a traducir, el primer aminoacil-ARNt (el ARNt cargado con el primer aminoácido), GTP (como fuente de energía) y factores de iniciación que ayudan a ensamblar el sistema de iniciación. La iniciación procariótica es el resultado de la asociación de las subunidades pequeña y grande del ribosoma y el acoplamiento del primer aminoacil-ARNt (fmet-ARNt) con el codón de iniciación mediante el emparejamiento de bases anticodón-codón.
El ribosoma consta de tres sitios: el sitio A (aminoacil), el sitio P (peptidil) y el sitio E (exit). El sitio A es el punto de entrada para el aminoacil-ARNt (excepto para el primer aminoacil-ARNt, fmet-ARNt, que entra en el sitio P). El sitio P es donde se forma el peptidil-ARNt. Y el sitio E es el sitio de salida del ARNt una vez descargado tras ofrecer su aminoácido a la cadena peptídica en crecimiento.  La iniciación termina cuando la subunidad ribosómica grande se une al sistema provocando el desacoplamiento de los factores de iniciación. Hay que tener en cuenta que las procariotas pueden distinguir entre un codón normal AUG (que codifica la metionina) y un codón de iniciación AUG (que codifica la formilmetionina e indica el comienzo de un nuevo proceso de traducción).
La iniciación termina cuando la subunidad ribosómica grande se une al sistema provocando el desacoplamiento de los factores de iniciación. Hay que tener en cuenta que las procariotas pueden distinguir entre un codón normal AUG (que codifica la metionina) y un codón de iniciación AUG (que codifica la formilmetionina e indica el comienzo de un nuevo proceso de traducción).
 La iniciación termina cuando la subunidad ribosómica grande se une al sistema provocando el desacoplamiento de los factores de iniciación. Hay que tener en cuenta que las procariotas pueden distinguir entre un codón normal AUG (que codifica la metionina) y un codón de iniciación AUG (que codifica la formilmetionina e indica el comienzo de un nuevo proceso de traducción).
La iniciación termina cuando la subunidad ribosómica grande se une al sistema provocando el desacoplamiento de los factores de iniciación. Hay que tener en cuenta que las procariotas pueden distinguir entre un codón normal AUG (que codifica la metionina) y un codón de iniciación AUG (que codifica la formilmetionina e indica el comienzo de un nuevo proceso de traducción).Elongación
La elongación de la cadena polipeptídica consiste en la adición de aminoácidos al extremo carboxilo de la cadena.
La elongación comienza cuando el fmet-ARNt entra en el sitio P, causando un cambio de conformación que abre el sitio A para que el nuevo aminoacil-ARNt se acople. Ahora el sitio P contiene el comienzo de la cadena peptídica de la proteína a codificar y el sitio A tiene el siguiente aminoácido que debe añadirse a la cadena peptídica. El polipéptido creciente que está conectado al  ARNt en el sitio P se desacopla del ARNt y se forma un enlace peptídico entre el último de los aminoácidos del polipéptido y el aminoácido que está acoplado al ARNt en el sitio A. En este punto, el sitio A ha formado un nuevo péptido, mientras que el sitio P tiene un ARNt descargado (ARNt sin aminoácido). En la fase final de la elongación, la traslación, el ribosoma se mueve 3 nucleótidos hacia el extremo 3' del ARNm. Como los ARNt están enlazados al ARNm mediante el emparejamiento de bases codón-anticodón, los ARNt se mueven respecto al ribosoma recibiendo el polipéptido naciente del sitio A al sitio P y moviendo el ARNt descargado al sitio E de salida. Este proceso está catalizado por un factor de elongación, gastando un GTP.
ARNt en el sitio P se desacopla del ARNt y se forma un enlace peptídico entre el último de los aminoácidos del polipéptido y el aminoácido que está acoplado al ARNt en el sitio A. En este punto, el sitio A ha formado un nuevo péptido, mientras que el sitio P tiene un ARNt descargado (ARNt sin aminoácido). En la fase final de la elongación, la traslación, el ribosoma se mueve 3 nucleótidos hacia el extremo 3' del ARNm. Como los ARNt están enlazados al ARNm mediante el emparejamiento de bases codón-anticodón, los ARNt se mueven respecto al ribosoma recibiendo el polipéptido naciente del sitio A al sitio P y moviendo el ARNt descargado al sitio E de salida. Este proceso está catalizado por un factor de elongación, gastando un GTP.
 ARNt en el sitio P se desacopla del ARNt y se forma un enlace peptídico entre el último de los aminoácidos del polipéptido y el aminoácido que está acoplado al ARNt en el sitio A. En este punto, el sitio A ha formado un nuevo péptido, mientras que el sitio P tiene un ARNt descargado (ARNt sin aminoácido). En la fase final de la elongación, la traslación, el ribosoma se mueve 3 nucleótidos hacia el extremo 3' del ARNm. Como los ARNt están enlazados al ARNm mediante el emparejamiento de bases codón-anticodón, los ARNt se mueven respecto al ribosoma recibiendo el polipéptido naciente del sitio A al sitio P y moviendo el ARNt descargado al sitio E de salida. Este proceso está catalizado por un factor de elongación, gastando un GTP.
ARNt en el sitio P se desacopla del ARNt y se forma un enlace peptídico entre el último de los aminoácidos del polipéptido y el aminoácido que está acoplado al ARNt en el sitio A. En este punto, el sitio A ha formado un nuevo péptido, mientras que el sitio P tiene un ARNt descargado (ARNt sin aminoácido). En la fase final de la elongación, la traslación, el ribosoma se mueve 3 nucleótidos hacia el extremo 3' del ARNm. Como los ARNt están enlazados al ARNm mediante el emparejamiento de bases codón-anticodón, los ARNt se mueven respecto al ribosoma recibiendo el polipéptido naciente del sitio A al sitio P y moviendo el ARNt descargado al sitio E de salida. Este proceso está catalizado por un factor de elongación, gastando un GTP.El ribosoma continúa trasladando los codones restantes del ARNm mientras siguen acoplándose más aminoacil-ARNt al sitio A, hasta que el ribosoma alcanza un codón de parada en el ARNm (UAA, UGA o UAG).

Terminación
La terminación ocurre cuando uno de los tres codones de terminación entra en el sitio A. Estos codones no son reconocidos por ningún ARNt. En cambio, son reconocidos por unas proteínas llamadas factores de liberación. Estos factores disparan la hidrólisis del enlace éster de la peptidil-ARNt y la liberación del ribosoma de la proteína recién sintetizada. O fin de la fase.Polisomas
La traducción es ejecutada por varios ribosomas al mismo tiempo. Debido al gran tamaño de los ribosomas, solo se pueden acoplar al ARNm a una distancia de 35 nucleótidos unos de otros. El sistema consistente en un ARNm y un cierto número de ribosomas se llama polisoma o poliribosoma.
Efecto de los antibióticos
Hay varios antibióticos que actúan interfiriendo en el proceso de traducción de las bacterias. Explotan las diferencias entre los mecanismos de traducción procariótica y eucariótica para inhibir selectivamente la síntesis de proteínas en las bacterias sin afectar al huésped. Algunos ejemplos incluyen:
La streptomicina provoca una mala lectura del código genético en las bacterias a concentraciones relativamente bajas e inhibe la iniciación a concentraciones mayores, enlazándose a la subunidad ribosómica 30s.
Las tetraciclinas bloquean el sitio A del ribosoma, evitando el acoplamiento de los aminoacil-ARNt.
El cloranfenicol bloquea la fase de la transferencia peptídica de la elongación en la subunidad ribosómica 50s, tanto en las bacterias como en las mitocondrias.



Hola me encanto tu blog! tiene información muy valiosa, tengo q hacer una exposición sobre OPERON DE LACTOSA ojala me puedas ayudar a entender mejor este término.
ResponderEliminarATTE: 1er semestre Univ. lorena Garnica
Ehh Mi tarea Dice Dibujar La traduccion Q Ocurre En Los Ribosomas Lo cuales Son Tres ARN'S
ResponderEliminarMi nombre es Nieves. Es que no encuentro otro modo de entrar y dejar mi comentario sin que sea por "anónimo", pero bueno, reiterarme en la calidad de este blog y darte las gracias.
ResponderEliminar